|
|
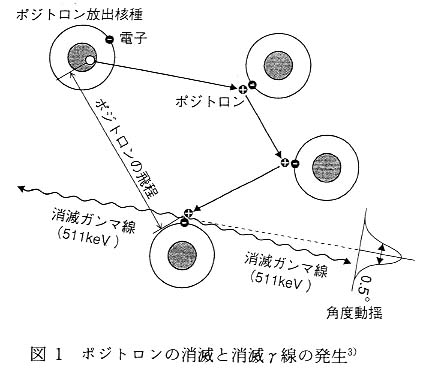
1. は じ め に 筆者らは, 悪化しつつある地球環境の中で食糧としての植物の生産を確保したり, 環境汚染物の除去や浄化技術の開発を最終的な目標として研究を行っている。 これらの達成のために必要となる知見は, 植物の環境変化に対する応答として植物機能にどのような変化が生じているかを詳細に調べることにより得られると考えた。 そして, 機能の変化による植物内の物質移行や分布変化を計測するための装置として植物用ポジトロンイメージング装置 (positron emit-ting tracer imaging system : PETIS) を日本原子力研究所と浜松ホトニクスの共同研究により開発した1)。 PETIS を用いたポジトロンイメージングでは, ポジトロン放出核種の崩壊により発生する消滅γ線を, 対向配置した二次元位置検出型検出器で同時計測することにより, 生きた植物中のポジトロン放出核種の経時的な二次元分布が画像として得られる。 現在は, 植物の機能を研究するために有用な種々のポジトロン放出核種で標識した化合物を製造し, これらの生きた植物内での移行, 蓄積, 分布等の計測およびその挙動解析を行っている。 これまでに整備できたポジトロンイメージング装置では, 植物の一個体すべてを計測視野内に納めることはできない。 そこで, 小型の植物であれば個体全部を同一視野に納めることができるイメージングプレート (imaging plate : IP) による計測を, 補助的な手法として利用している。 通常, 生物分野で利用されるトレーサといえば, 3H, 14C, 32P, 125I などの放射性同位元素や, 13C, 15N, 18O といった安定同位体が一般的であろう。 前号で渡辺らにより, なぜポジトロン放出核種を医学分野で利用するかが IP の応用例とともに紹介された2)。 本稿では植物, 特に個体としての植物を研究対象としている筆者らが, なぜポジトロン放出核種を選択したのか, ポジトロンイメージング計測のメリットはなにかをご理解いただくために, PETIS による計測と対比しながら, IP による植物ポジトロンイメージング計測について紹介する。 2. ポジトロン放出核種の利用 ポジトロン放出核種は, その名のとおりポジトロン (電子の反物質で陽電子とも呼ぶ, e+ または β+ と表す) を放出して β+ 崩壊するものの総称であり, 崩壊に伴って放射線を発生するいわゆるラジオアイソトープである。 筆者らがこれまでの研究で植物研究への応用を検討したポジトロン放出核種を表 1 に示す。 ここに挙げた核種のうち [22Na]NaCl は購入したものである。 また, [15O]H2O は半減期が 2 分と短く, 現在の高崎研究所のRI製造設備では供給までに時間がかかりすぎてしまうため, 浜松ホトニクスの中央研究所にある PET センターで製造し, 計測に供した。 崩壊に伴って放出されるポジトロンのエネルギーは単一ではない。 これは核から飛び出して周囲との相互作用によりそのエネルギーを徐々に落としながら, 最終的には電子と結合して消滅する。 このとき, 由来するポジトロン放出核種の種類やポジトロンが持っていたエネルギーにかかわらず, 511keV のエネルギーを持った 2 本の消滅γ線が同時に 180° 反対方向に発生する特徴を持つ (図 1 )。 筆者らが PETIS を用いて行っているポジトロンイメージング計測は, このポジトロンが消滅に伴って発生する 2 本の消滅γ線が同時に反対方向に飛ぶ特徴を利用している。 医学分野で応用されている PET (positron emission tomography : ポジトロン断層診断) も PETIS と同様にポジトロンの消滅に由来する消滅γ線を検出する計測法である。 PETIS については次の項で詳しく紹介することにして, ここではもう少しポジトロン放出核種とその標識化合物について述べることにする。
ポジトロン放出核種に由来する消滅γ線を検出対象とした計測法を, 個体レベルの植物研究に応用するメリットは次の 3 点に要約できる。 ① 生物の主要な構成要素である炭素[11C], 窒素 [13N], 酸素 [15O] が存在する。 ② 非破壊, 非接触での検出なので, 生きた植物内の情報を取り出せる。 ③ 半減期が短い核種([11C]: 20分, [13N] : 10分, [15O] : 2 分) では, 同一個体を用いた繰返し実験が可能である。 これらのメリットについてもう少し詳しく解説する。 まず, 1 番目のメリットについては, 植物の基礎的な代謝にかかわる重要な物質が, ポジトロン放出核種で標識した化合物として得られるということである。 実際には同じ計測装置により,[11C]-二酸化炭素,[13N]-硝酸,[13N]- アンモニア, [15O]-水といった標識化合物を用いた研究が可能である。 [11C]-二酸化炭素は言うまでもなく植物の特徴的な代謝活動のひとつである光合成の機能を調べる上で, きわめて有益である4)5)。 [13N]-硝酸, [13N]-アンモニアは窒素源の吸収・代謝からアミノ酸合成などに関する機能の研究に有効である6)-8)。また,[15O]水は植物に限らず生物の主成分であり, 特に植物では物質輸送の媒体として, また光合成の機能に蒸散という形で深く関与しているため, やはり有用な標識化合物である。 ここで [18F]F-イオンについて触れておきたい。 表 1 を見て [18F] だけが浮いたように感じられた方も多いのではないかと推察される。 フッ素は生体を構成する元素ではないし, むしろ異物ともいえる存在である。 これには [15O]-水の 2 分というあまりに短い半減期が関係している。 先にも述べたとおり, 現在筆者らが使用している施設では, [15O]-水の製造から供給までの所要時間の関係で計測開始時に十分な放射能を確保することができない。 しかし, 水の動きは植物の機能を調べる上で重要な要素であるので何とかして計測を行いたい。 そこで, 半減期が 110 分と長い [18F] は, 他の栄養成分のように代謝を受けたり特定の部位に蓄積あるいは分配される可能性が少なく, 水に極微量溶け込んでいる [18F]F-イオンは導管のような大きな水の通り道の中であれば水に準じた挙動を示すと考えられるので, 水の動きを示すひとつの指標として研究に供している9)。 また, [18F] は [11C] などとともに, 標識化合物の合成に用いる核種としても有用であり, 医学の領域では脳でのグルコースの代謝を調べるために [18F]FDG (2-フルオロデオキシグルコース) が PET 計測に供されている10)。 筆者らの研究においても, 植物による糖の移行や代謝を調べるために, [18F] で標識した [18F]FDG を合成し, 実験に供している。 ポジトロン放出核種を用いることの 2 番目のメリットは, 非破壊, 非接触での計測が可能なことである。 筆者らの研究では, 植物が環境変化に対してどのような応答を示すのかを, 植物内の物質の流れや分布の変化として捕らえようとしているのだから, 植物試料は生きた状態であることが望ましい。 この一方で, 必要な情報は植物試料の内部にあるのだから, 植物試料の厚さなどの影響を受けずにその情報を取り出す必要がある。 植物を対象としたトレーサについて考えてみると, [13C], [15N], [18O] などの安定同位体は, 植物の代謝機能の研究において貴重な知見をもたらしているが, 植物中にあるこれらの元素を計測するためには, 植物の切断や抽出といった操作が必要となり, 通常同じ植物を複数回の計測に供することは不可能である。 放射性同位元素について考えてみると,β崩壊により発生する放射線はα線,β線あるいはγ線であり, これらの放射線は物質透過力がそれぞれ異なる。α線の物質透過力はきわめて低く, 紙が 1 枚あっただけで止ってしまう。β線はこれより少し透過力は高いものの, 由来する核種によりエネルギーが異なるだけでなく分布を持っている。 したがって, 植物試料中から抜け出してくる 線を検出できても, 植物の厚さは部位によって異なり, 計測した値から植物内に 線を放出する核種がどれくらい存在していたかを算出することは困難である。 これに対してγ線は物質中での透過力が高いので, 試料の厚みやその不均一さの影響を受けることなく外部での検出が可能である。 したがって, 植物試料内から出てくるγ線を検出するような実験系が組めれば, 植物を磨り潰したり, 接触することなく計測することが可能となる。 PETIS によるポジトロンイメージング計測は, ポジトロンの消滅により発生する消滅γ線を検出する計測法であるから, 内部の情報を外から知ることが可能である。 植物に対して非破壊・非接触という計測環境を作り出すことは, 植物の環境を理想に近い状態に保つということだけではなく, 同じ植物個体を用いて複数回の実験が可能となる点で持つ意味は大きい。 これに関しては次の項にも関係があるので後述することにする。 ポジトロン放出核種に限って言えば, これらの核種が β+崩壊に伴い発生する消滅γ線は, 核種によらずすべてエネルギーが単一 (511keV) であるので, このエネルギーを持つγ線だけに絞っての計測が可能であり, 計測時のバックグラウンドのノイズを低減するという意味でも有利である。
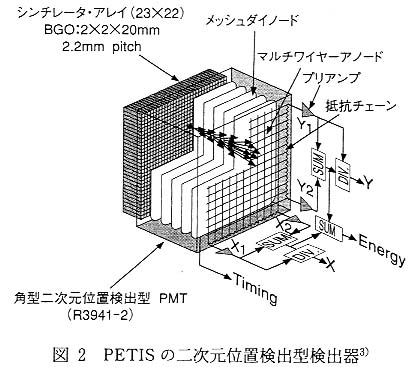
ポジトロン放出核種を用いる 3 番目のメリットとしては, 生体を構成する炭素, 窒素, 酸素といった元素に, 比較的半減期の短い核種が存在することである。 これらの元素についてトレーサとして利用されている核種についてみると, たとえば [14C] は歴史的な年代推定に使われていることからもわかるように, 半減期は 5730 年(3.0×109分)と [11C] の 20 分に比べ桁違いに長い。 このように半減期の長いトレーサを植物に吸収させると, 後々までそれが減衰せずに残るので, 冷却時間を多少とったとしても同じ植物を用いての複数回の実験はきわめて困難となる。 これに対してポジトロン放出核種では, [11C] は 20 分, [13N] は 10 分, [15O] は 2 分と非常に短い。 これらの核種で標識された化合物は, 植物に投与してから数時間を経過すれば植物中の放射能は検出限界以下まで減衰する。 したがって, 検出限界まで減衰すれば同一個体に対しての標識化合物の複数回投与が可能となる。 同じ植物を用いて異なった条件を設定して比較計測が可能になるのである。 また, 2 回目以降に使用する標識化合物が 1 回目の標識化合物と同じである必要はまったくない。 たとえば, [11C]CO2 を用いてまず光合成機能について計測を行い, 続いて [13N]NO3- や [13N]NH4+ を用いて窒素同化機能の計測をし, さらに [15O]H2O で蒸散機能を計測するというような, 従来のトレーサ実験では不可能な実験でさえ可能となる。 同じ植物を用いて複数回の実験が行えることで, 生物を用いた実験での最も厄介な問題とも言える個体差の影響を除くことが可能となる。 これは, 生物を対象とした研究を行うにあたっては大変大きなメリットとなる。 ポジトロン放出核種を用いる 3 番目のメリットとしては, 生体を構成する炭素, 窒素, 酸素といった元素に, 比較的半減期の短い核種が存在することである。 これらの元素についてトレーサとして利用されている核種についてみると, たとえば [14C] は歴史的な年代推定に使われていることからもわかるように, 半減期は 5730 年(3.0×109分)と [11C] の 20 分に比べ桁違いに長い。 このように半減期の長いトレーサを植物に吸収させると, 後々までそれが減衰せずに残るので, 冷却時間を多少とったとしても同じ植物を用いての複数回の実験はきわめて困難となる。 これに対してポジトロン放出核種では, [11C] は 20 分, [13N] は 10 分, [15O] は 2 分と非常に短い。 これらの核種で標識された化合物は, 植物に投与してから数時間を経過すれば植物中の放射能は検出限界以下まで減衰する。 したがって, 検出限界まで減衰すれば同一個体に対しての標識化合物の複数回投与が可能となる。 同じ植物を用いて異なった条件を設定して比較計測が可能になるのである。 また, 2 回目以降に使用する標識化合物が 1 回目の標識化合物と同じである必要はまったくない。 たとえば, [11C]CO2 を用いてまず光合成機能について計測を行い, 続いて [13N]NO3- や [13N]NH4+ を用いて窒素同化機能の計測をし, さらに [15O]H2O で蒸散機能を計測するというような, 従来のトレーサ実験では不可能な実験でさえ可能となる。 同じ植物を用いて複数回の実験が行えることで, 生物を用いた実験での最も厄介な問題とも言える個体差の影響を除くことが可能となる。 これは, 生物を対象とした研究を行うにあたっては大変大きなメリットとなる。 このように, ポジトロン放出核種を用いた計測では, その特徴を生かすことにより, これまでのトレーサ実験法では得られなかったような知見が十分期待できるのである。 それでは, ポジトロン放出核種を用いたイメージングの実際について以下に紹介する。 3. 植物研究用ポジトロンイメージング装置 筆者らがポジトロンイメージング計測に使用している PETIS は, 計測対象となる小型の植物試料には厚みのある部分が少ないことから, 医学分野で実用化されている PET のように立体物の断層画像を得るためのものではなく, 二次元での投影画像を得るための装置として開発を行った。 医学分野でのポジトロンイメージング技術の利用に関しては, 前号の渡辺の解説2)や, PET 計測技術や臨床応用を紹介した文献10)11)があるので, 参照していただきたい。 PETIS の計測原理1)は, 一対の対向配置した二次元位置検出型検出器から等距離面上に配置した植物試料中のポジトロン放出核種に由来する一対の消滅γ線を同時計測し, それぞれの検出器への入射位置を直線で結んだ中点をポジトロンの消滅位置とするものである。 PETIS を用いた計測では, 数多く生じる消滅γ線の中から対となる消滅γ線を特定するために, 一方の検出器で検出したγ線の入射に対して, もう一方の検出器でのγ線の検出にタイムウインドウを設定している。 PETIS の検出器に用いている検出用素子は, 2mm×2mm×20mm の Bi4Ge3O12 シンチレータで, これを 2.2mmピッチで 22×23 個並べたシンチレータアレイを角型二次元位置検出型光電子増倍管に装着してある (図 2 )3)。 1 台の検出器は 48mm×50mm の計測視野を持つ。 小型の植物試料が対象とはいえ, この計測視野は情報を得るためには十分とは言えない。 そこで, PETIS では対となる検出器を増設することにより, 視野が拡張できるように設計されている。 現有の PETIS では 3 対の検出器を搭載し約 5cm×25cm の視野が得られるが, さらに 3 対の検出器を増設することで, 約 15cm×25cm の視野が得られるよう装置の拡張を進めている。
植物ポジトロンイメージングに使用しているもうひとつの計測装置が, IP に蓄積した放射線エネルギーに関する情報の読出しと画像解析を行うバイオイメージングアナライザ (BAS, 富士写真フイルム) である。 計測では対象となるラジオアイソトープがトリチウムかそれ以外の核種かにより, 使用する IP が異なる。 筆者らはポジトロン放出核種をトレーサとして用いているので, 汎用タイプで大きさが 20cm×40cm の IP を計測に使用している。 IP からの情報の読出しには MacBAS1500 を使用しており, 読取り画素サイズは 100m が得られる。 ちなみに, 20cm×40cm の IP が使用でき, BAS1500 の後継機種である BAS2500 では, 読取り画素サイズは 50m に向上している。 4. ポジトロンイメージングの実際 筆者らが植物中のポジトロン標識化合物の移行・分布計測の実験を行うときの流れを図 3 に示す。 まず, 試験目的に従い植物試料を調製し, 計測する部位を PETIS の視野に収まるようセッティングを行う。 計測に使用するポジトロン標識化合物を製造し, 必要に応じて精製等の処理を行う。 植物にポジトロン標識化合物を投与し, PETIS による計測を開始する。 計測終了後, 植物試料内のポジトロン標識化合物に残っている放射能が減衰するのを待って, 繰り返して計測を行う場合には標識化合物を製造するステップに戻る。 これ以上計測を行わない場合には, 植物試料全体の最終的な標識化合物の分布状況を知るために, BAS を用いた IP 計測を行う。 以下に PETIS および IP 計測の例を示しながら, ポジトロンイメージング計測について紹介する。
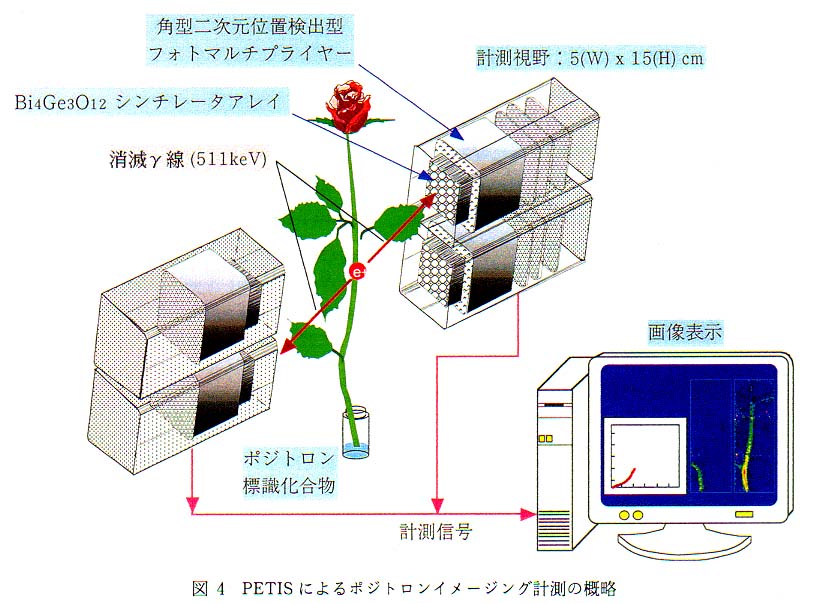
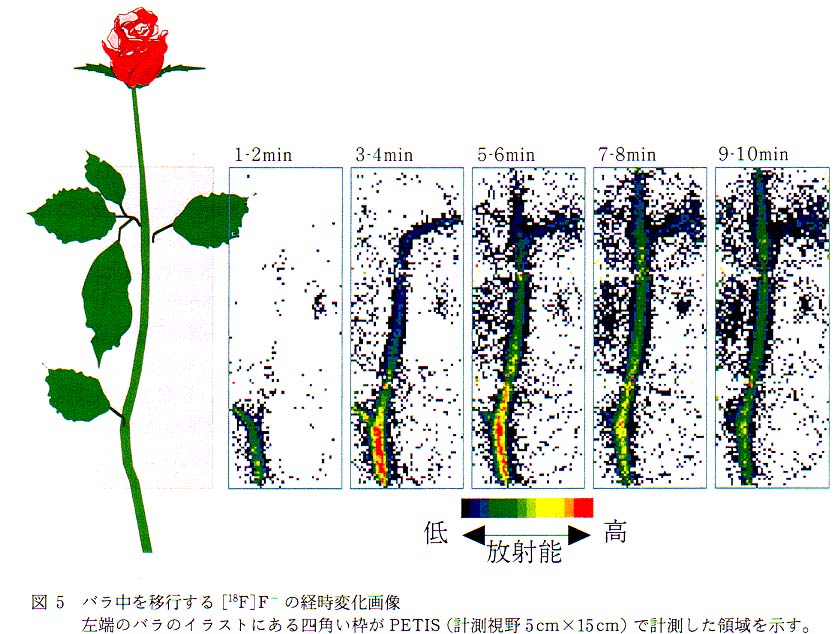
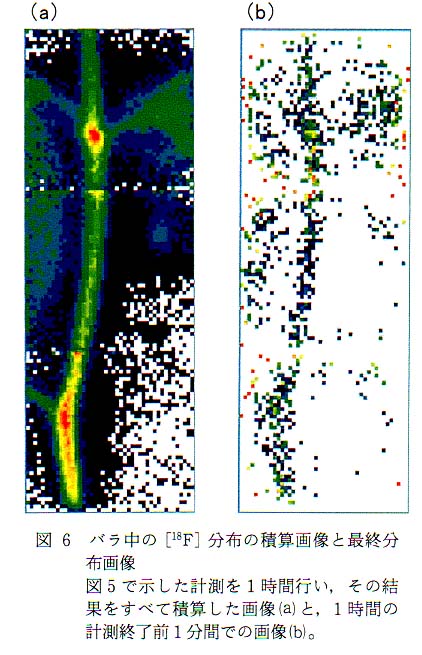
図 4 は検出器を 2 台搭載した PETIS (計測視野 : 5cm×15cm)により,バラ中での[18F]F- イオンの移行計測の概略である。 茎より投与した [18F]F- イオンは導管流にのってバラの中を移動する。 PETIS の計測視野内で発生した消滅γ線は対向配置した検出器で同時計測され, その信号は位置演算回路で処理後パーソナルコンピュータに送られ, 一定時間ごとの [18F]F- イオン分布画像を逐次表示するとともにデータとして蓄積される。 PETIS で計測信号の処理から再構築画像の表示までに要する時間は, 主にコンピュータの処理速度に依存し, 現有装置は 15 秒に 1 回画像を更新する性能を持つ。 通常筆者らが行っている計測では, 植物内での標識化合物の移動の速さと, 鮮明な画像を得るためのデータ蓄積との兼ね合いから, ほとんどの計測が 30 秒から 1 分で画像を更新する設定を用いている。 図 5 にバラ中を移行する [18F] の 1 分間おきの分布変化画像例を示す。 このように PETIS 計測では, 植物中の物質移行や分布の経時変化を秒から分のオーダーで可視化できる点が, 従来のトレーサを用いた計測法にはない特徴である。 さて, PETIS 計測では最後に更新された画像が植物中での最終的な標識化合物の分布を示すものであり, 計測をとおしての積算画像とは異なる。 計測の開始から終了までに蓄積されたデータを使って積算画像を再構築すると, 図 6 に示したような画像が得られる。 積算時間が長くなればなるほど 1 画像あたりの検出γ線量は増えるので, 再構築された画像は見やすいものとなるが, [18F] の移動過程も積算しているので, 最終的な [18F] の分布とは異なったものとなる。 最終的な[18F] の分布といった場合には図 6 に示した画像がこれに相当する。
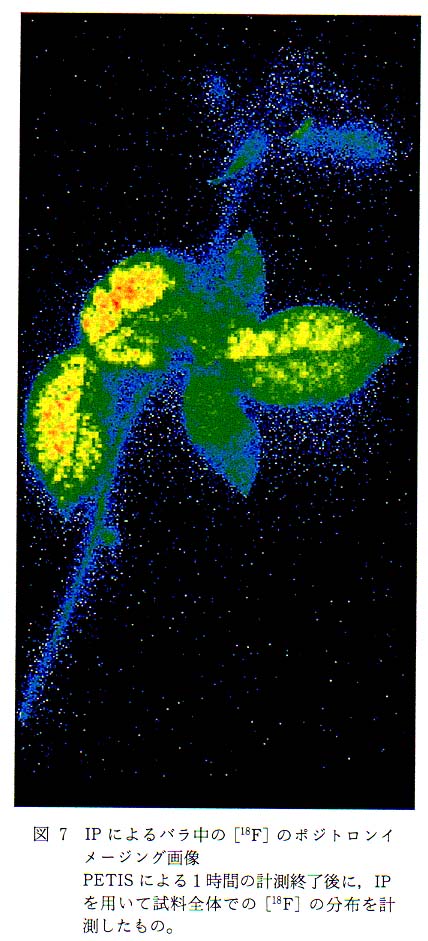
計測に用いたバラは茎丈が約 30cm なので, PETIS の計測視野ではすべてをカバーすることはできない。 そこで, PETIS での計測終了後, 全体への標識化合物の移行状態を確認するために,IP を用いて計測を行う。図 7 がPETIS計測終了後の IP を用いた BAS 計測で得られた画像である。 バラのような試料は当然のことながら, 葉は薄く平面的な組織なのでプレートと密着させやすいが, 茎はその曲がり具合によってはプレートとどうしても接触しない部分が出てきてしまう。 また, 花の部分は茎よりさらに厚みがあるため, その付け根はプレートから浮いてしまい, 花たくや花びらも一部分しかプレートに接触させることができない。 筆者らが計測に使用している MacBAS1500 の解像度は 100m である。 したがって, PETIS より一桁高い解像度が得られることになる。 ところが, 試料とプレートを密着させることができないと, この高解像度を十分生かすことができない。 植物個体が試料の場合, きれいな切片を作成すること自体が困難である上, 作業の過程で試料中の放射能は減衰してしまう。 したがって, BAS の性能・特徴を生かしきるための試料調製は, 短寿命の核種を用いてのポジトロンイメージングでは難しい。
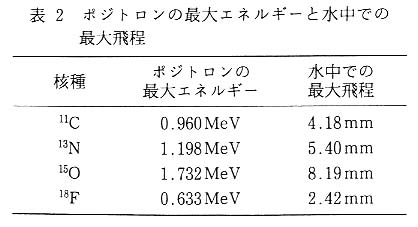
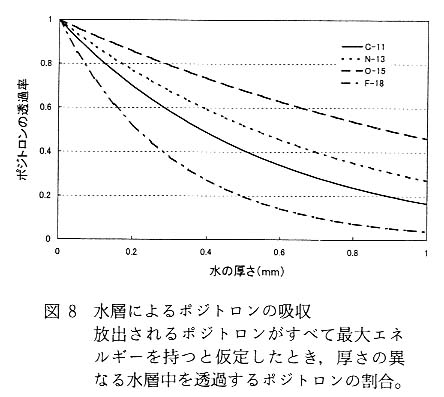
さて, 図 6 と図 7 は [18F]F- イオンをバラに投与してから 1 時間後の [18F] の分布を示しているのだが, 両者を比較すると, PETIS の画像では [18F] は茎の部分に多く存在し, 葉の部分に少ないことがわかる。 一方, IP 画像では茎より葉に [18F] が多く分布しているように見られる。 図 7 に示した場合に限らず IP 画像では, 葉が最も鮮明に現れ [18F] の分布も多い画像が得られる。 これは, 上述のとおり葉はプレートに密着させることができることが一因と考えられるが, もうひとつの要因として, 検出している放射線の種類が考えられる。 PETIS 計測で用いるポジトロン放出核種は, 崩壊様式がβ+ だけの核種でも,γ線 (ポジトロン) および消滅γ線を発する。 このほかの崩壊様式をあわせ持つものでは, さらに放射線の構成とそのエネルギー分布は複雑になる。 [18F] は, β+崩壊が 96.7%, EC 崩壊が 3.3% である。 したがって[18F]の崩壊に伴って発生する放射線は, エネルギー分布を持ったβ線, 511keV のγ線およびごく少量の特性X線ということになる。 PETIS では消滅γ線を選択的に検出するような計測を行うが, IP では放射線に対する選択性はない。 まず,β線が持つエネルギー分布と植物試料などによる吸収との関係について説明する。 表 2 に筆者らが計測に多用しているポジトロン放出核種に由来するβ線 (ポジトロン) の最大エネルギーと水中での最大飛程を示した。 それぞれの核種についてポジトロンがすべて最大エネルギーを持つとして, 水中での透過率を計算により求めると図 8 のようになる。 ここでは, 水の厚さが 1mm までの透過率を示したが, たとえば [18F] に由来するポジトロンの最大エネルギーは 634keV で, このエネルギーを持ったポジトロンが完全に吸収されるためには水層として 2.42mm が必要となる。 図 8 に戻り [18F] に由来するポジトロンの吸収曲線を見ると, 1mm 厚の水があるとこれを透過できるポジトロンはわずか数%となる。 これは, あくまでもポジトロンすべてが最大エネルギーを持っていると仮定したときの値であり, 実際にはポジトロンのエネルギーには分布があるので, さらに低い値となる。 一般にポジトロンが持つエネルギーの分布中心は, 最大エネルギーのおよそ 13 付近にあるとされており, 筆者らが行ったポジトロンの吸収効率計測試験の結果でも, これを裏付けるような結果が得られており,[18F] に由来するポジトロンはわずか 1mm 厚の水が存在するだけでほぼ完全に吸収されてしまい, 外には出てこないことになる12)。 したがって, IP を用いたバラの計測では, 肉厚の薄い葉ではプレートに到達するポジトロンの割合が, 茎に比べて多くなることは容易に想像でき, 見かけ上葉に存在する [18F] が茎に存在する [18F] より多くなると考えられる。
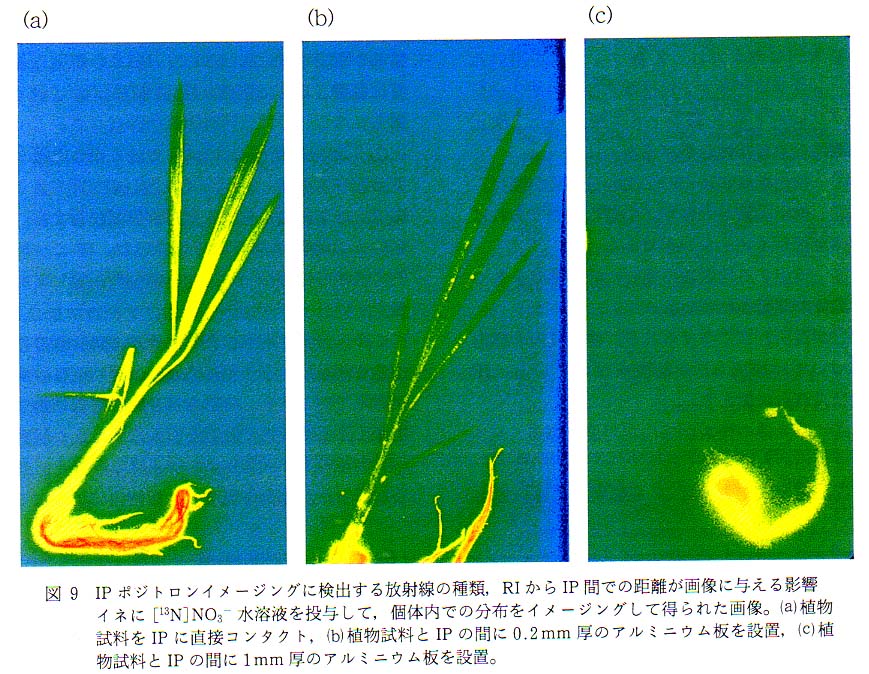
上記のこと, および消滅γ線の発生位置が画像に与える影響をモデル実験により確認した結果が図 9 の画像である。 ポジトロン標識化合物として [13N]NO3- 水溶液を吸収させた植物試料 (イネ) と IP との間にアルミニウム板を置いてイメージングを行った。 植物試料を直接 IP にコンタクトさせた画像では, 経根吸収された 13NO3- が根に多く蓄積されているものの, すべての葉の先端にまで分配されている様子が観察でき, 展開葉については葉脈による硝酸濃度の違いについても観察可能であった。 また, 根についても込み合っていない部分については 1 本ごとの識別も可能であった。 植物試料と IP との間に 0.2mm 厚のアルミニウム板を挟んだときの画像では, 全体の色調が低放射能側にスライドしてはいるものの, 葉先までの画像が得られた。 また, 得られた画像上の位置分解性も遮蔽体がない場合とほとんど変化がないように見られた。 これに対して植物試料と IP の間に 1mm 厚のアルミニウム板を設置すると, 根の部分に放射能が検出できたが, 茎や葉については画像がまったく得られなかった。 また, 根の部分についても 1 本ごとの根を確認することはできず, 全体にぼやけた画像となった。 0.2mm 厚のアルミニウム板の面積密度は 0.54gcm2 であり, 表 2 に示したように [13N] 由来のポジトロンの水中最大飛程は 5.4mm であるので, すべてのポジトロンが IP に到達せずにアルミニウム板中で消滅していることになる。 したがって, 図 9 と との差が, ポジトロンによる作像への寄与分を表すことになる。 さて, 図 9 と は, ポジトロンが画像に与える影響を取り除き, 消滅γ線だけでイメージングを行うために植物試料と IP との間に挟んだアルミニウム板の厚さが異なるだけで, その他の計測条件は同じであるが, 図 9 は葉脈や根などの細かい部分が観察できるのに対し, 図 9 では地上部は観察することができず, 根の部分も全体がぼやけていて, 根の 1 本 1 本を見ることができなかった。 このような結果になった理由は, 植物試料の IP からの距離にあると考えられる。 わずか 1mm の距離と感じられる方も多いかと思われるが, 消滅γ線はポジトロン放出核種の位置から 360° 確率的に発せられる。 ポジトロンイメージング装置を用いた定量計測のために, 試料とポジトロン吸収体との距離がイメージの質に与える影響を評価した実験結果では, ポジトロンの消滅位置, すなわち消滅γ線の発生位置が焦点面から 1mm 離れた場合でも画像の劣化は生じた。 したがって, 読取り画素サイズが小さい IP 計測では, その影響が顕著に現れることが容易に想像される12)。 また, 試料の厚さをきちんと評価することも難しいのが現状である。
これまでの経験や実験の結果をまとめると, 厚さが比較的均一で薄い試料であれば IP を用いたポジトロンイメージングにより, 植物内の標識化合物の分布を比較的よく反映した画像が得られるが, 厚さが不均一でプレートに密着させにくい試料では, 量的な解析や評価を行う場合に注意が必要となる。 IP の放射線のエネルギーや線種に対する特性については, 文献を参照していただきたい13)-18)。
生きた植物のイメージング計測は, 植物の分子生物的な手法により得られた植物の生体活動に伴う現象が, 個体レベルの植物内でどのように機能しているかを探るための重要な手がかりを与えてくれる。 イメージングプレートは画像化計測法として, 他の計測法にはない魅力的ともいえる特徴を持っている。 このような特徴を生かした計測が数多く行われ, 計測手法として改良・確立されていくことにより, 植物分野の研究進展に一層寄与することを期待しつつ, 本稿を終わりたい。
|