|
|
**大阪バイオサイエンス研究所神経科学 565-0874 大阪府吹田市古江台 6-2-4 ***京都大学大学院情報学研究科知能情報学専攻 606-8501 京都市左京区吉田本町
Development and Application of in vitro Positron-Emitting Tracer(PET) Method. Yasuyoshi WATANABE*,**, Fusao NAKAMURA**, Masaaki TANAKA** and KiyoshiMATSUMURA**,***: *Department of System Neuroscience, Osaka City University Graduate School of Medicine, 1-4-3 Asahimachi, Abeno-ku, Osaka 545-8585, Japan, **Department of Neuroscience, Osaka Bioscience Institute, 6-2-4 Furuedai, Suita-shi, Osaka 565-0874, Japan, ***Department of Intelligence Science and Technology, Kyoto University Graduate School of Informatics, Yoshida-honmachi, Sakyo-ku, Kyoto 606-8501, Japan.
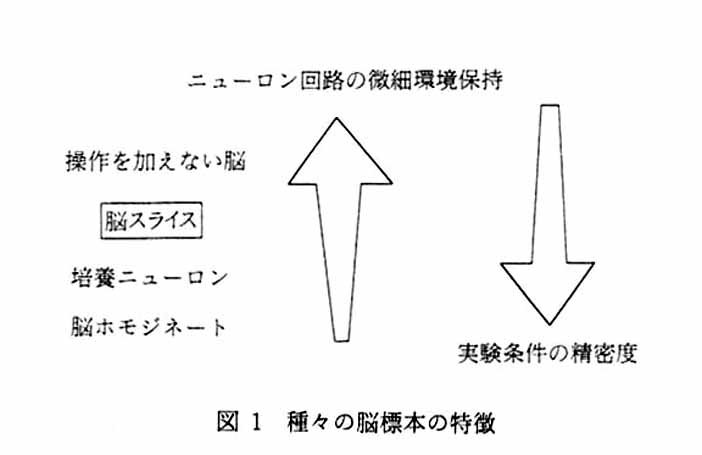
筆者らは, 1993年から1997年の 5 年間, 科学技術振興事業団の国際共同研究プロジェクトの一つとして, スウェーデン・ウプサラ大学 PET センターと 「サブフェムトモルバイオ認識プロジェクト」 を行った。 ポジトロンエミッショントモグラフィ (PET) の基礎・応用技術に関わる技術革新と超高感度インビボ生化学計測がテーマの柱であった。 プロジェクトでのさまざまな技術革新の中で, 新規設計合成トレーサ動態を PET のように個体丸ごとで計測する系に進めるまでの路程の中で, 通常のインビトロ計測へのポジトロン核種の利用やインビボ-インビトロの中間段階の計測法を行って, トレーサの挙動や可否を検討してきた。 そのような計測手段の一つとして, 「インビトロ PET 法」 (この場合, PET は, Positron-Emitting Tracer と読む1)。) あるいは, 最近, 「バイオラジオグラフィー」 (BF 研究所所長 澤田 徹先生の命名) と名付けている方法を創始した。 本法により, 機能・代謝として生きている臓器スライスでの二次元分子イメージングが可能になったので, 利用の仕方によって, 非常に価値ある発明であると思われる。 日本研究者のオリジナリティが生かされている発明で, 今のところ, ウプサラ大学 PET センター附属 JST ラボ (前臨床ラボ), 大阪市立大学医学部 RI 実験施設, 福井医科大学高エネルギー医学研究センター, 放射線医学総合研究所, 東京都老人総合研究所などで, 行われている実験方法であるが, すでに, 外国医薬品企業や他のポジトロン核種にアクセスできる研究所では, 組織培養系や術後腫瘍塊を用いた同様の技術が使われている。 この方法の創造には, 全身オートラジオグラフィ (ARG) やラジオルミノグラフィ (RLG) の技術が欠くべからざるものである。 1960-1970年代にかけてのウプサラ大学故 Ullberg 教授らの全身 ARG の開発は, クリオスタット, 乳剤, トリチウム感受性フィルムの開発など, さまざまな面でこのような技術の発展に寄与してきたが, その歴史の中で, 実は, ポジトロン核種である 11C と他の長半減期核種とのダブルラベリング法を最初に始めたのもウプサラ大学 PET グループとこのグループなのである2)。 起源はここにあると言っていい。 筆者らは, 1993年以前にも, 何度か生きて機能している脳スライスなどでこのポジトロン核種を使って二次元的トポロジーを探ろうというアイディアを討論していたが, 実際には富士写真フイルムによって RLG の技術が確立されるまでは, 実現を見ていない。 結局, ここで述べるインビトロ PET 法 (バイオラジオグラフィ) の技術は, エネルギーが高く薄いポリエチレンフィルム (サランラップなど) を突き抜ける β+ 放出核種 (ポジトロン核種) と高感度でダイナミックレンジの広いイメージングプレートがなければ, 達成されていない技術なのである。 先人に心より感謝する。 筆者らのプロジェクトでは, 基本的にポジトロン核種とイメージングプレートを使う方法論について, 数々の基礎的実験を行った3)4)。 2. インビトロ PET 法の実際 ここでのインビトロ PET 法には, 脳スライスを用いる。 脳機能の研究には正常な脳に始まり, 単離した培養神経細胞に至る種々の脳標本が用いられている (図 1 )。 脳機能の解明という観点から見ると, それぞれの標本は固有の長所短所をあわせ持つ。 丸ごとの脳は, その機能, 神経細胞の結合等すべてが保たれているが, 精緻な実験的操作は困難である。 一方, 単離した神経細胞では実験的操作は容易であるが, 神経結合は失われており脳機能の本質を担う機能的神経ネットワークに迫ることはできない。 脳スライス標本はこれらの中間に位置する。 すなわち, 局所的に神経結合は保たれており, また血液脳関門もなく実験的操作も比較的精緻に行うことができる。 このような特徴を持つ脳スライス標本は脳機能研究において広く用いられてきた。
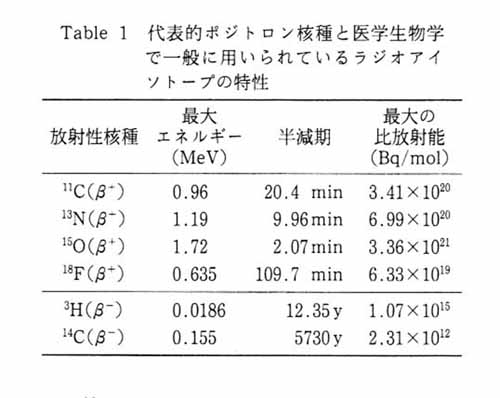
しかし, これまで脳スライスの生化学的過程のイメージングはまったく行われていない。 これが実現すれば今まで PET のために開発されてきた種々のトレーサを有効に活用して, 精密度の高いインビトロの実験を行うことが可能になり, また新規 PET トレーサを開発する上で有用な基礎的データを提供することができる。 何故ポジトロン核種なのか?という基本的なポイントを理解してほしい。 表 1 に代表的なポジトロン核種と医学生物学で一般に用いられているラジオアイソトープ核種の特性を比較した。 第一に, ポジトロン核種から放出されるポジトロンは, トリチウム (3H) 等の β- の持つエネルギーレベルと比べて高エネルギーである。 この性質は, 厚みのある生体組織や, 培養液存在下で信号を検出することに有利である。 第二に, ポジトロン核種の半減期は短いものが多い。 もし同じモル数の分子がアイソトープで標識されているとすると, 14C あるいは 3H が数十年かけて出すカウントを, ポジトロン核種は数分で出すことを意味する。 つまり, 理論的な比放射能が高いということである。 したがって, ポジトロン核種を用いることにより, 分の単位でイメージングに必要な信号を得ることができ, スライスを生かしたままで経時的なイメージングが行える可能性がある。 さらに, 時間をおけば, 同一サンプルで一つの標識化合物のイメージングを行ったあと, 異なった種類のトレーサで違う情報を得るという, 複数のトレーサを用いたマルチトレーサ実験が可能になる。 これらのことはポジトロン核種でのみ実現しうる。
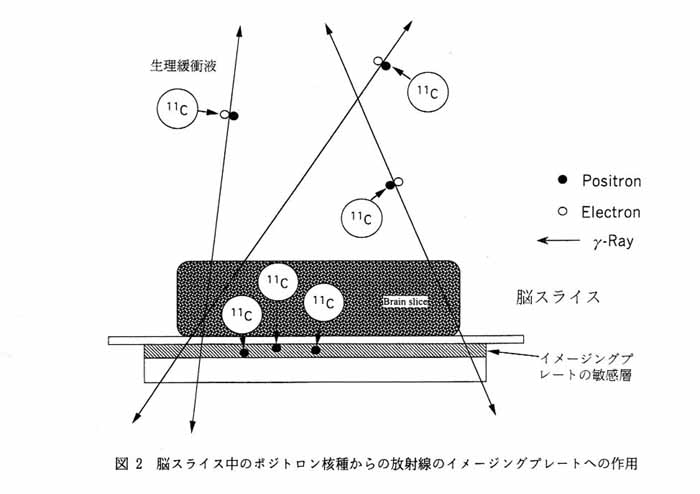
図 2 にインビトロ PET の原理を示す。 脳 (組織) スライスが十分量の酸素 (95% O2/5% N2 混合ガス) で飽和された生理緩衝液中に置かれている。 スライスと薄膜 (約 30 ミクロン厚) を隔ててイメージングプレートが配置されている。 この緩衝液にポジトロン核種で標識したトレーサを加えると, トレーサは脳スライスに存在する特異的受容体に結合するか, あるいは細胞内に取り込まれ代謝を受ける。 このようにしてスライスに蓄積したポジトロン核種は, その物理学的崩壊に従ってポジトロンを放出する。 イメージングプレートの近傍から放出されたポジトロンはイメージングプレートに衝突して信号を残すが, イメージングプレートから遠い部位で放出されたポジトロンは電子と衝突した後, 消失しγ線となる。 このγ線の大部分はイメージングプレートを貫通し, そこに信号を残さない。 したがって, この方法ではスライスに集積したポジトロン核種の分布を選択的に検出し, 背景にある溶液中の非結合状態のポジトロン核種からの影響を少なくする。 従来の ARG 法ではフリーのトレーサを洗い去った後にのみ, イメージングが可能であったが, ポジトロン核種とイメージングプレートを用いるとフリーのトレーサ存在下での組織内のイメージングが可能となる。 さらに先に述べたようにポジトロン核種を用いると, 短時間 (数分) のうちに画像化に必要な信号が得られる。 したがって, フリーのトレーサ存在下で同一スライスに集積してくるトレーサの分布を経時的にイメージングすることが可能となる。
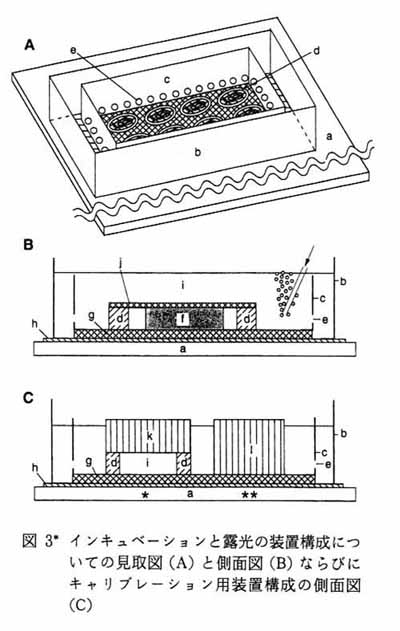
図 3 にインビトロ PET 実験装置の概略を示す。 実験装置はプラスチック製の二つのチャンバで構成される。 外チャンバはその底面がポリプロピレンあるいはサランラップ等の薄膜で構成される。 外チャンバは緩衝液を満たすため, 水漏れのないようにすることが重要である。 これらの薄膜とポリスチレン製の外枠の接着には防水性両面テープを用いる。 内チャンバは外チャンバに入れることのできる大きさのもので, 底面はナイロンネットで構成される。 実際にはナイロンストッキングを強く均一に伸展させ, エポキシ接着剤でポリスチレン製の枠に接着している。 内チャンバの側面には緩衝液の流入を容易にするための孔が一定間隔であけられている。 内チャンバを緩衝液で満たした外チャンバに入れ, ナイロンネット上に脳スライス標本 (200 - 300μm 厚) を置く。 スライスの浮遊を防ぐためにステンレス性リングの上面にナイロンネットを接着した外枠を用いる。 ステンレスリングの厚みはスライスと同じであり, このリングをスライスにかぶせることにより, スライスを圧迫することなくその場に保持することができる。 スライスに集積したトレーサの定量化のための較正用セットアップを図 3C に示す。 スライスと同じ厚みの既知の濃度 (N nM) のトレーサ溶液層を図のように形成し, そのイメージングプレート上でのカウント (A) とバックグラウンドのイメージングプレート上でのカウント (B) を求める。 さらに実際のスライスのある領域のカウント (C) とバックグラウンドのカウント (D) を求める。 スライスのある領域でのトレーサ濃度 (X nM) は以下の式で算出する。 X=N×(C/D)/(A/B) この式が成立するのは放射能量とイメージングプレートのカウントが直線関係にある範囲であり, 目安として 20 分間の曝露でイメージングプレートのカウントが 10 以上の場合とした。
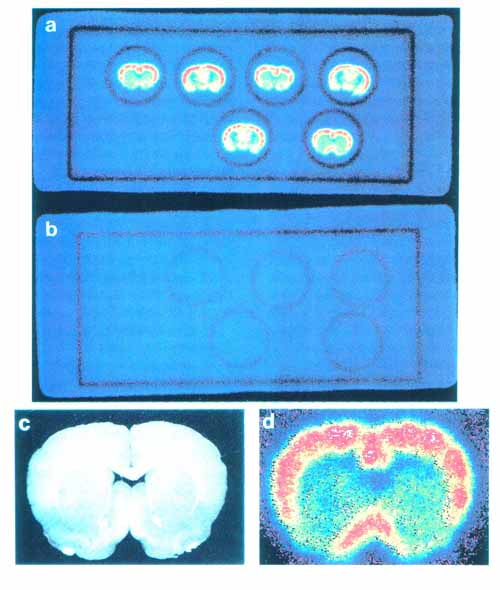
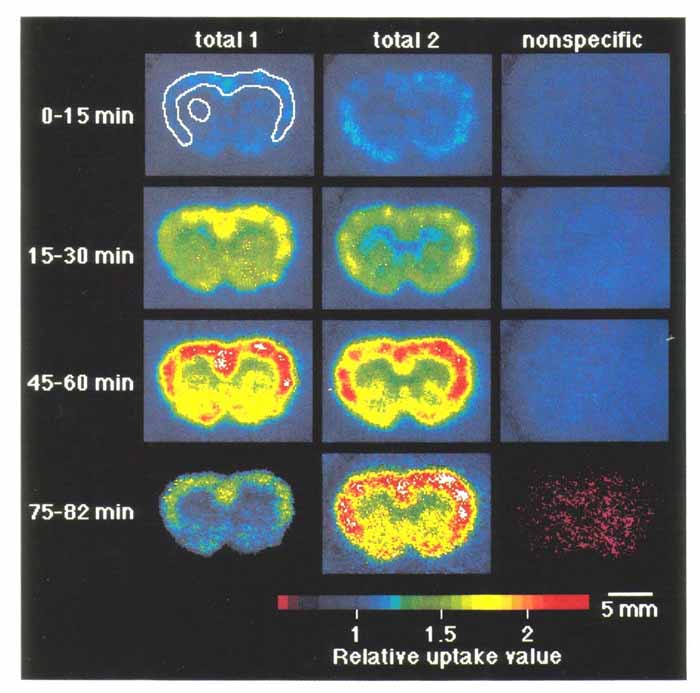
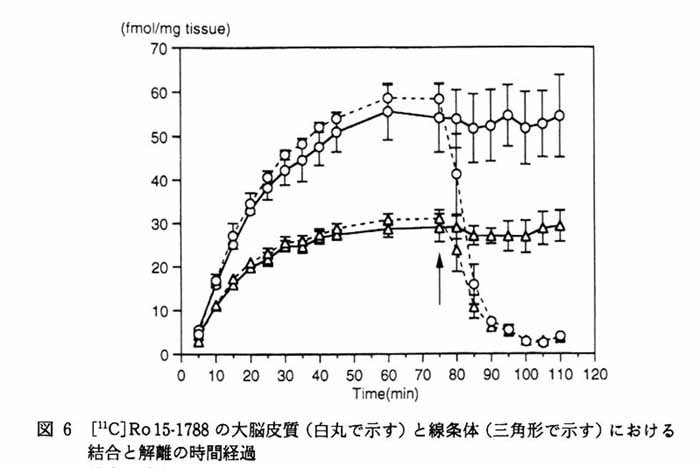
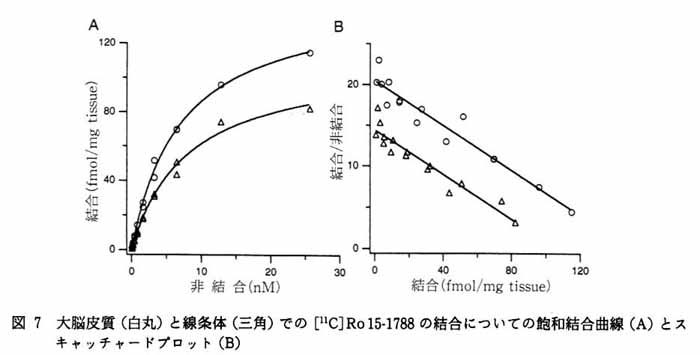
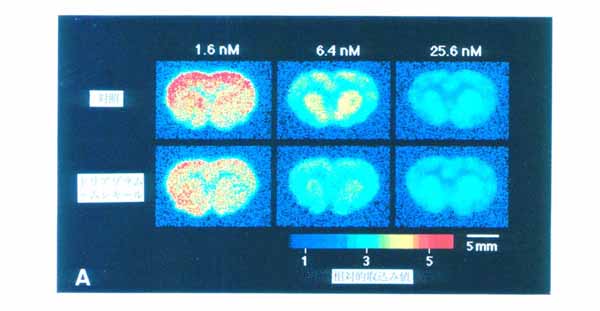
3. 本方法の応用 3・1 脳スライスにおける受容体結合の イメージング5) 図4にベンゾジアゼピン受容体アンタゴニストである [11C]Ro15-1788 の脳スライスへの結合実験の画像を示す。 a は 5nM の [11C]Ro15-1788 を加えて 60 分後の画像で, total binding を表している。 一方 b は a と同じ量のトレーサとともに 1μM の非標識 Ro15-1788 を加えたもので, 非特異的結合を表している。 すなわち画像 a と b の差が特異的結合になる。 c は脳スライスの実際の像, d はそのスライスで得られた[11C]Ro15-1788 結合の拡大図である。 大脳皮質, および前脳基底部に高密度の結合が存在することが明らかである。 図 5 は 15 分の時間間隔で [11C]Ro15-1788 の集積経過を同一スライスで記録したものである。 図 6 はスライスへの [11C]Ro15-1788 の集積を時間軸に沿って定量的に示している。 大脳皮質, 線条体ともに 60 分間のインキュベーションで平衡状態に達している。 またこの状態で高濃度の非標識 Ro15-1788 を加えるとその結合が低下していく経過が観察された。 図 7 は異なった濃度の [11C]Ro15-1788 を用いてスキャチャードプロット解析を行った結果である。 大脳皮質, 線条体とも解離定数は 7nM となった。 最大結合部位量は大脳皮質で 146fmol/mg tissue, 線条体で 107fmol/mg tissue となった。 このようにインビトロ PET 法により生きている脳スライスにおいて, 受容体のイメージングとその定量的解析が可能となった。 この手法は他の受容体リガンドに対しても適用可能である。
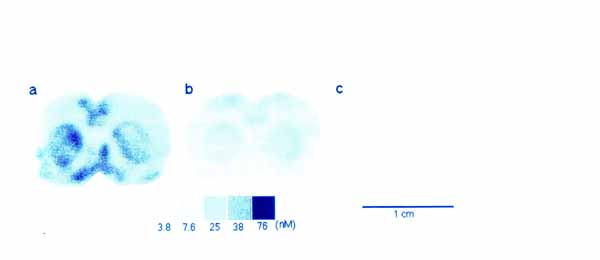
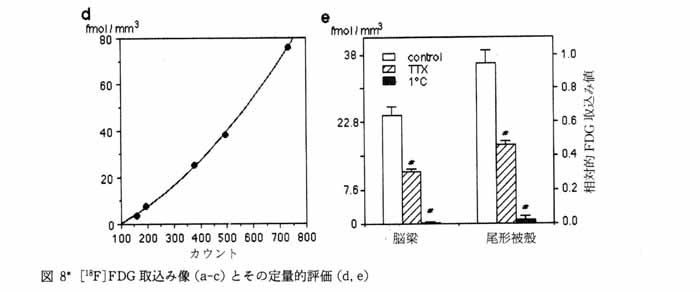
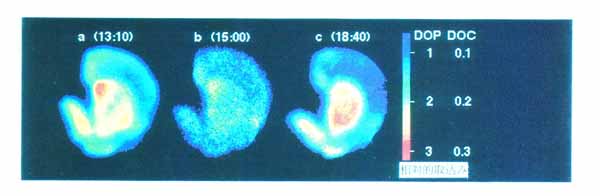
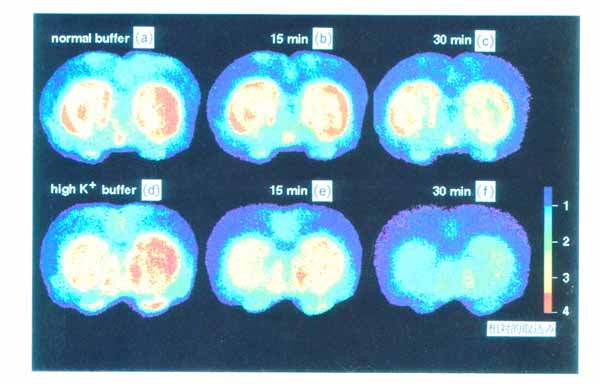
3・2 脳スライスにおける代謝のイメージ ング1) ブドウ糖代謝のトレーサとして PET で広く用いられている [18F]FDG をスライスを含む緩衝液中に投与し 37 ℃ で 45 分間放置し, その後 30分 間通常の緩衝液で洗う。 これにより解糖系の律速酵素 (ヘキソキナーゼ) の活性が画像として得られる (図 8a)。 培養温度を 1 ℃ にした場合は [18F]FDG の取込みは激減 (図 8c) し, また電位依存性ナトリウムチャンネルの阻害剤であるテトロドトキシン (TTX) 存在下で 37 ℃ で培養した場合にも有意にその取込みが抑えられる (図 8b)。 このことは脳スライスへの [18F]FDG の取込みがスライスの糖代謝を反映しており, また神経活動に伴う神経およびグリアの糖代謝を TTX により識別できることを示す。 神経伝達物質のドーパミンの前駆体である L-[11C]DOPA を緩衝液中に投与して, 線条体を含む脳スライスを培養した。 その結果, ドーパミン神経の終末が豊富に存在する線条体で高濃度の取込みが見られた(図 9 )。 このイメージが L-[11C]DOPA そのものの存在を表しているのか, あるいは L-[11C]DOPA が dopa-decarboxylase によって転換された [11C]dopamine を表しているのか明らかにするために, カルボキシル基 11C 標識の L-DOPA(DOC) と側鎖 β-位 11C 標識の L-DOPA(DOP) を用いた。 DOC は dopa-decarboxylase で代謝されると 11C 標識-二酸化炭素として脳外に速やかに排出されるが, DOP は dopamine になった後も放射能を保持する。 図 9 に示すように, DOP で培養したときのみ線条体に高濃度の放射能の取込みが見られた。 したがって, DOP 存在下で培養した際に見られる線条体の放射能は [11C]dopamine の量を反映していると考えられる。 さらに [11C]dopamine の放出も経時的にイメージングが可能である。 特に, 脱分極刺激として高カリウム緩衝液中で洗うと放出が促進されることも観察された (図10)。 以上のようにインビトロ PET 法では, 糖代謝のみでなく, アミノ酸代謝と神経伝達物質放出のイメージングが可能となった。
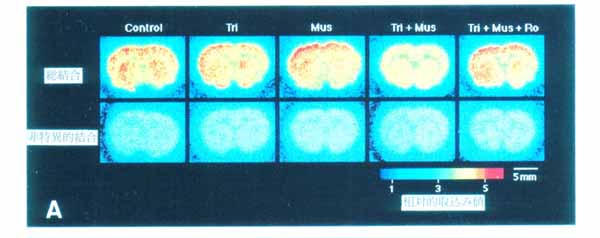
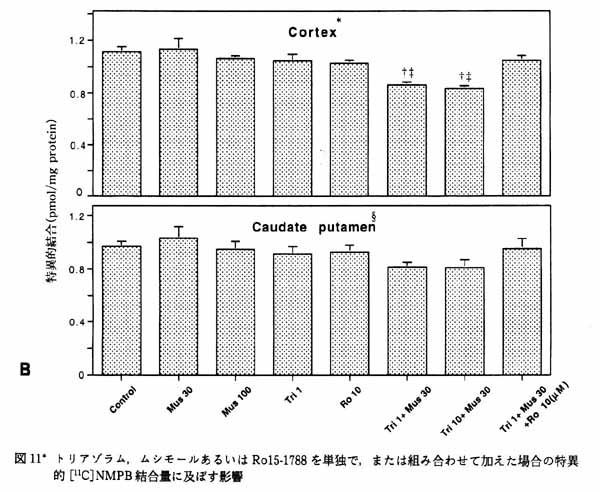
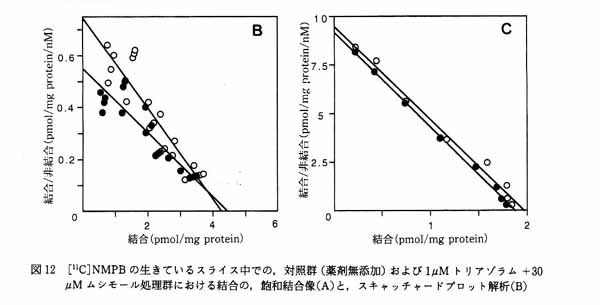
3・3 ムスカリン受容体結合に対する GABA/benzodiazepine の作用6) 睡眠導入薬として用いられているトリアゾラム (ハルシオン) は, benzodiazepine(Bz) 受容体にアゴニストとして作用し, GABA の作用を増強する。 臨床的にはトリアゾラムに順向性健忘の副作用があることが報告されている。 一方, 脳内ムスカリン性アセチルコリン系が記憶に関与しているということが知られている。 したがって, トリアゾラムがムスカリン性アセチルコリンシステムに何らかの影響を与えている可能性が考えられた。 この点に関して 11C 標識 N-methyl-piperidyl-benzilate ([11C]NMPB ; ムスカリン性受容体アンタゴニスト) の脳スライスへの結合を指標に検討した。 [11C]NMPB の線条体, 大脳皮質への結合は, トリアゾラムと muscimol (GABAA 受容体アゴニスト) 共存下で低下した (図11)。 この低下はベンゾジアゼピン受容体アンタゴニストである Ro15-1788 で消失することから, ベンゾジアゼピン受容体を介したものと思われる。 Scatchard plot 解析によりこの低下は, 受容体とリガンドの結合の親和性が低下したことを反映していることが明らかとなった (図12A と B)。 一方, 従来から一般的に用いられてきた, 細胞膜分画における結合実験ではこのようなトリアゾラムと muscimol による協奏作用は見られなかった (図12C)。 このことは細胞が生きていること, あるいは受容体周囲の何らかの微小環境がトリアゾラムの作用発現に必要であることを示唆する。 さらにスライスと細胞膜分画では [11C]NMPB 結合の親和性が大きく異なり, 後者が前者の 20 倍の値になる。 この差違が何に起因するのかは現在不明である。 このように脳スライス標本では細胞膜分画では観察できない現象の研究を可能にする場合もある。 生きている脳により近い, 脳スライスにおける研究の重要性が示唆された。
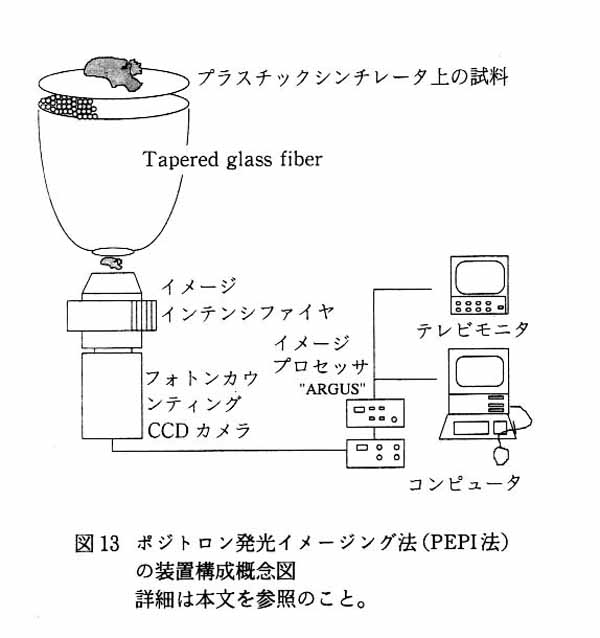
4. まとめと将来展望 本法を用いて, 生きている脳スライスにおけるさまざまな生化学過程の経時的イメージングに世界で初めて成功した。 前述のように, この成功は高エネルギー, 短半減期というポジトロン核種の特性と RLG 技術を生かしたことに拠るところが大きい。 脳スライスを用いることの意義は, 局所的神経回路が保持されている, 実験的操作が容易である, 細胞が生きている, という点である。 特に細胞が生きているという点は, 代謝性トレーサを用いたトータルの代謝実験には不可欠であり, すりつぶした脳サンプルや, 凍結切片ではこの種の実験は不可能である。 また受容体結合実験は脳の膜分画や凍結切片でも可能であるが, 今回の実験で示したように脳スライスでなければ観察できない微細環境による相互作用がある。 これは, PET データ, 実際の脳機能の分子神経機序を理解する上で, 非常に重要な点で, 従来の方法論ではアプローチできなかった世界である。 本法の適用範囲はきわめて広い。 今後の発展方向としてはサンプルの多様化, トレーサの多様化, 刺激方法の多様化, 時間・空間分解能の向上という三つの方向性が考えられる。 サンプルの多様化としては今回は長期培養を考えない実験系の脳スライスを用いたため, 同一サンプルでの繰返し実験が 12 時間程度に限られたが, スライス培養法を用いれば数日間にわたって同一サンプルでの実験が可能であろう。 脳の発生・発達, シナプス発達などの研究にも標的分子の追跡の系として, これからの発展が期待される。 脳の共培養系を用いて神経回路の形成過程で受容体・代謝系がどのように変化するか, という問題に本手法はきわめて効果的であろう。 さらにサンプルは脳に限らず, 生体内臓器のすべてが対象となりうる。 トレーサの多様化としてはアンチセンスオリゴヌクレオチドの導入が挙げられる。 そのためには半減期の比較的長いポジトロン核種 (76Br, 半減期 16 時間, など) を用いて, スライス培養系で実験を行うことが望まれる。 刺激方法の多様化としては局所的な電気刺激の導入, 細胞内セカンドメッセンジャー系特異的な薬物刺激, アンチセンスオリゴヌクレオチドの導入が考えられる。 空間・時間分解能は現在使用している RLG システムに大きく依存する。 このバイオラジオグラフィ法は, 一度に多数のスライス標本からの記録ができる利点があるが, ① 1 画像を得るためには 10 - 20 分の記録時間が必要で, 短時間の薬理学的な反応に追従できない点, ②電気生理学的な刺激や記録を併用するためには, 1 画像ごとにイメージングプレートの交換操作があるためにサンプル側を動かさなければならない, などの欠点がある。 この点を補うために, 筆者らは新しいインビトロ PET 法として, Positron Emission Photon Imaging 法 (PEPI 法)7) を開発した。 これは, アクリルチャンバーの底に薄いプラスチックシンチレータを貼り付け, シンチレータ上の脳スライスが取り込んだポジトロン核種からのポジトロンが発光させた微量発光を, 光ファイバによってイメージインテンシファイヤに導き, フォトンカウンティング CCD カメラとイメージプロセッサ (ARGUS 50, 浜松ホトニクス) によって画像を構築するものである (図13)。 これによって得られる時間分解能はバイオラジオグラフィ法に比してかなり改善され, 今のところ 2 分ごとの脳スライスイメージングができるようになった。 今後, 種々のアイソトープで標識した物質の代謝をこの時間分解能で画像化できるものと期待される。
謝 辞 最後に, この原稿を書く機会を与えていただいた栗原紀夫先生の暖かいご支援と福井医科大学の村田哲人先生, 米倉教授はじめ, ウプサラ大学のロングストレーム教授など多くの共同研究者に感謝いたします。
文献 |